

Différence entre les protistes et les champignons

- 803
- 27
- Carla Lefevre
Introduction
Les protistes et les champignons comprennent deux royaumes uniques de la vie. Les protistes démontrent une variance solide des traits compliquant leur taxonomie. Les champignons sont beaucoup plus simples à caractériser. Les champignons divergeaient des protistes environ 1.Il y a 5 milliards d'années [1] un événement qui a entraîné une perte de flagellum lors de la transition d'un habitat aquatique à terrestre avec le développement concomitant de nouveaux mécanismes de dispersion de spores non aquatiques [2] [2]. Les champignons ont adapté un ensemble de caractéristiques reconnaissables qui aident à clarifier leurs différences par rapport aux protistes.
Différences cellulaires
Les protistes sont des organismes unicellulaires. La majorité des champignons sont multicellulaires et structurés dans un système filamenteux allongé de ramification des hyphes [1]. Les structures hyphes ramifiées sont constituées d'une ou (généralement) de cellules supplémentaires enfermées dans une paroi cellulaire tubulaire [1]. La plupart des protistes sont de forme sphérique, qui est sous-optimale pour obtenir de l'oxygène par diffusion. Les grands protistes ont une forme allongée pour répondre à leur besoin accru de diffusion d'oxygène [3].
Taille de la cellule
Les protistes unicellulaires sont pour la plupart microscopiques mais des exemples rares ont été trouvés des milliers de mètres carrés dans la zone [3]. Les champignons sont généralement suffisamment grands pour être observés à l'œil nu, mais un grand nombre d'espèces microscopiques existent [1].
Membrane cellulaire
Les protistes peuvent contenir des parois cellulaires de type végétal, des parois cellulaires en forme d'animaux et même des pellicules offrant une protection contre l'environnement extérieur [3]. De nombreux protistes n'ont pas de paroi cellulaire [3]. Contrairement à la variété de membrane cellulaire protiste, une caractéristique déterminante des champignons est la présence omniprésente d'une paroi cellulaire chitineuse [14].
Organisation intracellulaire
Les champignons sont composés d'un système alambiqué de compartimentation des hyphes par un système de partitionnement de septa [1]. Les septa n'ont été trouvés dans aucun protist [3]. Les septa fongiques divisent les hyphes en compartiments perméables [1]. La perforation des septa permet de transloquer les organites, y compris les ribosomes, les mitochondries et les noyaux entre les cellules [3]. Les organites protiste existent dans un cytoplasme non compartimenté [3].
Appendices cellulaires
Contrairement aux champignons principalement stationnaires, les protistes sont mobiles [1,3] et cette motilité différencie les protistes morphologiquement des champignons par l'ajout d'appendices cellulaires. Les protistes contiennent fréquemment des appendices comme les cils, les flagelles et les pseudopodes [3]. Les champignons n'ont généralement pas d'appendices cellulaires bien que de rares exemples d'appendices conidaux dans des champignons existent [4].
Respiration
Respiration protiste
i) Respiration aérobie protiste
Les protistes obtiennent l'oxygène par diffusion et cela limite leur capacité de croissance cellulaire [3]. Certains protistes comme les phytoflagellates effectuent un métabolisme hétérotrophique autotrophique et oxydatif [3] [3]. Le métabolisme protiste fonctionne de manière optimale à travers une large gamme de températures et de quantités de consommation d'oxygène. Il s'agit d'un sous-produit de la pléthore de niches qu'ils habitent, qui ont une vaste gamme de températures et de disponibilité en oxygène [3].
ii) Respiration anaérobie protiste
La respiration anaérobie obligatoire existe parmi les protistes parasites, une rareté pour les eucaryotes [3]. De nombreux protistes anaérobes obligatoires manquent de cytochrome oxydase entraînant des mitochondries atypiques [3].
iii) Respiration fongique
La plupart des champignons respirent de manière aérobie en utilisant des chaînes respiratoires ramifiées pour transférer des électrons de NADH à l'oxygène [5]. Les NADH déshydrogénases fongiques sont utilisées pour catalyser l'oxydation de la matrice NADH et sont capables de le faire même en présence de certains inhibiteurs comme la roténone [5]. Les champignons utilisent également d'autres oxydases pour respirer en présence d'inhibiteurs pour l'ubiquinol: la cytochrome c odoréductase et la cytochrome c oxydase [5]. Les oxydases alternatives permettent probablement une pathogénicité efficace en présence de mécanismes de défense de l'hôte à base d'oxyde nitrique [5].
Osmorégulation
Les protistes qui habitent un environnement aqueux ont une amplification des structures cellulaires que l'on ne trouve pas dans les champignons. Cette amplification permet un degré d'osmorégulation plus élevé. Les vacuoles contractiles sont des organites protistes qui permettent l'osmorégulation et empêchent l'enflure et la rupture cellulaire [3]. Les vacuoles contractiles sont entourées d'un système de tubules et de vésicules appelé collectivement le spongiome qui aide à l'expulsion des vacuoles contractiles de la cellule [3]. Les vacuoles contractiles sont nettement moins abondantes dans les champignons [1,3].
Différences mitochondriales
Génomes mitochondriaux provisibles
Contrairement aux champignons, les génomes mitochondriaux provistiques (MT) ont conservé un certain nombre d'éléments génomiques proto-mitochondrians ancestraux. Cela est évident par la réduction des gènes des champignons mtgénomes [6]. Les mtgénomes provistiques varient en taille du génome de 6 kb de Plasmodium falciparum au génome de 77 Ko du choanoflagellate Monosiga brevicollis, Une gamme plus petite que les champignons [6]. La taille moyenne de la mtgénome provistique est de 40 Ko significativement plus petite que la taille du génome mitochondrial fongique moyen [6].
Les mtgénomes provisibles sont compacts, riches en exon et souvent composés de régions de codage qui se chevauchent [6]. L'espace intronique non codant représente moins de 10% de la taille totale de la taille du protiste [6] [6]. Une grande partie de l'ADNmt protiste n'a pas d'introns du groupe I ou du groupe II [6]. La teneur en A + T est plus élevée dans les mtgénomes provisibles par rapport aux champignons [6].La teneur en gènes des mtgénomes protiste ressemble plus aux mtgénomes végétaux que les mtgénomes fongiques [6]. Contrairement aux champignons, les mtgénomes protistes codent pour les ARN de grande et petite sous-unité [6].
Génomes mitochondriaux fongiques
Les champignons ont évolué à partir des protistes et leur divergence se caractérise par la réduction des gènes et l'addition d'intron [6]. Par rapport aux mtgénomes protistes riches en gènes, les mtgénomes fongiques contiennent une pléthore de régions intergéniques composées de répétitions et d'introns non codantes qui sont principalement des introns du groupe I [7]. La variation de la taille des mtgénomes fongiques s'explique principalement par les régions intron plutôt que par la variance basée sur les gènes trouvées dans les mtgénomes provistiques [7]. Les régions intergéniques représentent jusqu'à 5 Ko de longueur dans les mtgénomes fongiques [7].
Bien que les mtgénomes provistiques contiennent plus de gènes, les mtgénomes fongiques contiennent une quantité significativement plus grande de gènes codants d'ARNt [6,7]. Les tailles de mtgénome fongiques s'étendent sur une plus grande gamme par rapport aux mtgénomes protistes. Le plus petit mtgenome fongique connu est de 19 kbp, trouvé dans Schizosaccharomyces pombe [6]. Le plus grand mtgenome fongique connu est de 100 kbp, trouvé dans Podospora Anserina [6]. Contrairement aux mtgénomes protistes, la teneur en gènes de l'ADNmt fongique est relativement cohérente entre les organismes [6].
Sources en nutriments et stratégies d'acquisition des nutriments
Acquisition de nutriments de champignons
Les champignons utilisent le mycélium, leur collection d'hyphes, pour acquérir et transporter des nutriments à travers la membrane plasmique de leurs cellules [2]. Ce processus dépend fortement du pH de l'environnement à partir duquel les nutriments sont acquis [2]. Les champignons sont des saprotrophes, acquérant leurs nutriments principalement à partir de la matière organique dissoute de décomposition des plantes et des animaux morts [1]. Toute digestion requise des nutriments se produit extracellulaire par la libération d'enzymes qui décomposent les nutriments en monomères à ingérer par diffusion facilité [1]
Acquisition de nutriments provisibles
Les protistes, en revanche, obtiennent leurs nutriments à travers une variété de stratégies. Une tentative de catégoriser les stratégies d'acquisition de nutriments protiste définit six catégories [3]:
- Producteurs primaires photo-autotrophes - Utilisez la lumière du soleil pour synthétiser les nutriments de CO2 et H2O.
- Baciti et détritivores - Se nourrir de bactéries ou de détritus.
- Saprotrophes - Nourrir de la matière sans vie extracellulaire et absorbée par extracellulaire.
- Algivores - Nourrir principalement des algues.
- Omnivores non sélectifs - Nourrir de manière non sélective sur les algues, les détritus et les bactéries.
- Prédateurs de Raptoriels - Nourrir principalement des protozoaires et des organismes de niveaux trophiques plus élevés.
Beaucoup de stratégies susmentionnées sont mixotrophiques. Par exemple, les producteurs primaires photoautotrophes comprennent des organismes marins qui peuvent utiliser différents niveaux d'hétérotrophie permettant une acquisition de nutriments qui ne nécessite pas d'apport d'énergie de la lumière du soleil lorsque la lumière du soleil n'est pas disponible [3].
Différences de reproduction
Les protistes et les champignons comprennent tous deux des espèces qui se reproduisent sexuellement et. Les protistes sont uniques en ce qu'ils incluent des organismes capables de reproduction aesexuelle et sexuelle au cours de la même vie [8]. La complexité de certains cycles de vie protiste entraîne des variations morphologiques étonnantes au sein de la vie de l'organisme qui permet des méthodes de reproduction distinctes [8]. Les changements morphologiques liés à la reproduction ne sont pas observés dans les champignons dans la mesure où ils sont chez les protistes.
Différences de reproduction aesexuelles
La reproduction aesexuelle dans les champignons se produit par la détente de spores émanant des corps fruitiers trouvés sur le mycélium ou par la fragmentation du mycélium ou par un bourgeon [9]. La reproduction aesexuelle chez les protistes se produit à travers une variété de méthodes. La fission binaire (division nucléaire unique) et la fission multiple (divisions nucléaires multiples) sont deux méthodes de reproduction aesexuelles communes parmi les protistes [8]. Une autre stratégie de reproduction spécifique au protiste est la plasmotomie [8]. La plasmotomie se produit parmi les protistes multinucléés et implique une division cytoplasmique sans division nucléaire [8].
Différences de reproduction sexuelle
La reproduction sexuelle est plus couramment mise en œuvre par les champignons [8,9]. Il est également plus complexe que la reproduction aesxuelle et nécessite donc une description plus détaillée pour établir une compréhension de la façon dont le processus diffère entre les protistes et les champignons.
Reproduction sexuelle fongique
Pendant la reproduction sexuelle fongique, la membrane nucléaire et le nucléole (généralement) restent intacts tout au long du processus [9]. La plasmogamie, la caryogamie et la méiose comprennent les trois étapes séquentielles de la reproduction sexuelle fongique [9]. La plasmogamie implique une fusion protoplasmique entre les cellules d'accouplement qui amène les noyaux haploïdes distincts dans la même cellule [9]. La fusion de ces noyaux haploïdes et la formation d'un noyau diploïde se produisent au stade de la catégorie [9]. Vers la fin de la caryogamie, un zygote existe et la méiose se déroule par la formation de fibres de fuseau dans le noyau. Cela rétablit l'état haploïde par séparation du chromosome diploïde [9].
Les stratégies fongiques pour l'interaction des noyaux haploïdes pendant la reproduction sexuelle sont plus variées en champignons par rapport aux protistes. Ces stratégies incluent la formation de gamètes et la libération de Gametangia (organes sexuels), l'interaction de gamétanges entre deux organismes et l'interaction des hyphes somatiques [9].
Reproduction sexuelle provisoire
Les stratégies de reproduction sexuelle protiste sont presque entièrement différentes de celles employées par des champignons. Ces stratégies impliquent des processus uniques qui diffèrent en raison de la structure cellulaire, en particulier des appendices cellulaires disponibles pour le contact avec d'autres protistes [8]. La formation et la libération de gamètes sont une méthode de reproduction sexuelle parmi les protistes flagellés très mobiles [8]. La conjugaison est une méthode utilisée par les protistes ciliés qui implique la fusion de noyaux gamtiques plutôt que la formation et la libération de gamètes indépendants [8]. L'autogamie, un processus d'auto-fertilisation qui est toujours considéré comme une forme de reproduction sexuelle, produit l'homozygotie parmi la progéniture d'une cellule parent auto-fertilisée [8].
Table
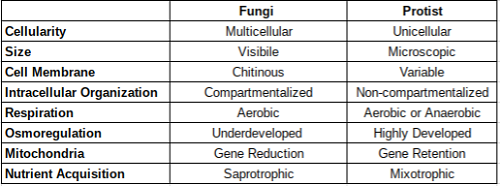
Comme résumé ci-dessus, les différences entre les protistes et les champignons sont vastes et peuvent être observées à tous les niveaux de structure et tout au long de leurs interactions comportementales avec leur environnement. Cette revue n'est qu'un résumé des différences. Les références citées fournissent des explications plus approfondies pour ceux qui souhaitent en savoir plus.